2025-03-13 10:30:08
天生的方向感——鸟类的指南针是怎么工作的?
从“木落雁南度,北风江上寒”到“万里人南去,三春雁北飞”,鸿雁南渡北归的身影穿梭于千年前的诗词文脉里,是古人对候鸟长距离迁徙的描述与惊叹;无论是汉代苏武牧羊中鸿雁传书的典故,还是唐代张九龄宰相飞鸽传信的轶事,都反映出鸟类在另一种归巢任务中同样精准的定向能力。
其实,强大的定向导航能力不只见于大雁和鸽子,而是许多鸟类都具备的特质:每年,数十亿只小型鸣禽离开北极和温带繁殖区,跨越数千公里的距离在热带和亚热带地区越冬。鸟类当中不乏卓越的远航者:斑尾塍鹬 (Limosa lapponica)从阿拉斯加飞往新西兰的旅程持续 7-9 天,从不停歇;北极燕鸥 (Sterna paradisaea) 在北极附近繁殖,在南极附近越冬,其航程跨越整个星球;还有海鸥,它们每年飞行超过十万公里,只为返回浩瀚海洋中的小岛繁衍生息[1]……从古至今,无论个例还是广例,无不展现着鸟类导航能力的强大。
图一 斑尾塍鹬 (Limosa lapponica)
图二 北极燕鸥 (Sterna paradisaea)
图源网络
面对如此强大的定位能力,人类固有的优越感荡然无存。同样是脊椎动物,共享同样的视听觉感受机制,甚至拥有不及人类拳头大小的脑,鸟类天生的方向感究竟从何而来?除了更高分辨率的常规感知之外,唯一可能的理由,就是它们具备人类不配备的特殊感觉系统。
那么,到底是什么特殊的地球物理量蕴含着精度极高的位置和方向信息,服务于鸟类的远程飞行?经过数个世纪的(de)研(yán)究(jiū),科(kē)学(xué)家最终把目光锁定在一类特殊的物理量——地磁场——之上。
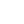
地磁场是地球表面及近地空间中存在的天然磁场。如图,磁场的强度由磁力线的疏密表征,在两极地区最大,在赤道(dào)地区最小。磁场形状类似条形磁铁的偶极子场,磁场的方向可以由磁力线方向表示,从南半球出发,跨越地球表面,在北半球重新进入地球内部。磁力线和地球之间形成的倾角随纬度而变化——在磁赤道处,磁力线平行于地球表面,倾角为 0°;从磁赤道向两侧,倾角逐渐陡峭,磁极处磁力线直接向下指向地球,倾角为 90°[2]。场强和倾角均可以编码位置信息——由于不同区域的磁场强度存在差异,因此前者可起到“地图”的作用;后者则携带方向信息作为“指南针”。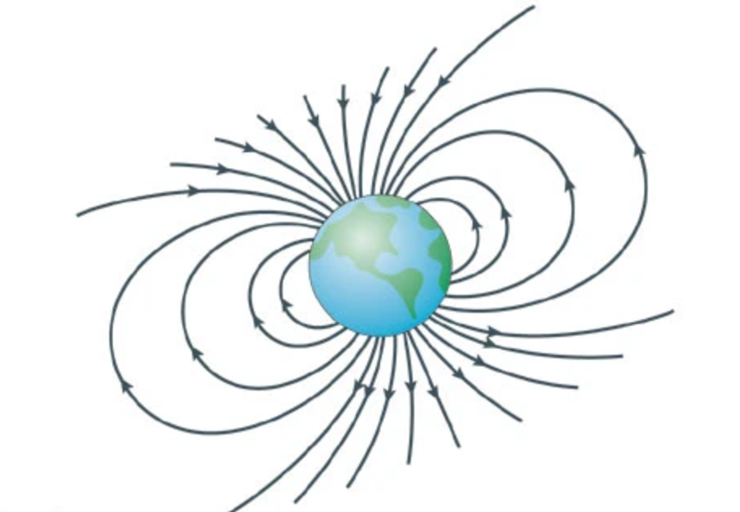
图三 地球磁场[2]
对于鸟类利用磁场导航的可能性早在19世纪就被提出和讨论。在Alexander von Middendorf关于俄罗斯鸟类的论文中指出:“......候鸟惊人的一心一意——不顾风和天气,不顾黑夜和雾——可能是因为家禽总是知道磁极的方向,因此也知道如何准确地保持它的方向”[3]。
动物感知地磁场能力的存在这一观点具有革命性意义,但直到20世纪中期才具备充分的技术条件加以检验。20世纪50年代,通过外加亥姆霍兹线圈干扰定向迁徙的行为学实验,科学家首先证明了一种夜间迁徙的鸣禽,欧洲知更鸟(Erithacus rubecula),具备磁场导航的能力[4]。
在随后的几十年里,绝大部分鸟类的磁场导航能力都得到了验证;有意思的一点是,鸟类磁定向系统需要依赖光照发挥作用:在可透弱光的木箱中长距离运输鸽子,随后记录其归巢方向选择,发现当在木箱周围施加随机干扰磁场时,鸽子的确完全失去了定位能力;而将箱子完全隔绝光照,也能起到与干扰磁场一致的效果,证明鸽子的磁定向能力依赖于光照的存在,即使是很弱的光强[5]。
磁感受依赖光照的特殊性质,意味着鸟类似乎能“看”到磁场;换言之,视觉系统与磁场感知密切相关。然而,磁场信息是如何经历一系列物理化学变化转化为神经信号的?近一个世纪以来,经过无数的假设与检验,鸟类磁感觉的神秘面纱正在被一点点揭开。
目前领域内主流的观点认为,磁场的强度和方向分别被两个正交的感觉系统编码:前者会通过激活鸟类头部含有磁铁矿晶体的感受器细胞,使鸟类感知磁场强度进而得出位置信息,与光照关联不大;后者则通过视觉系统起作用,以一种奇妙而罕见的自由基对机制影响视网膜上的化学反应方向,最终使鸟类得以感知磁场方向。在此我们将主要探讨后一种机制——鸟类是如何“看”到磁场方向的?
自由基对参与鸟类磁场感知的理论可以追溯到 1978 年,Schulten 等提出磁敏感光化学转变的假说,自由基对作为反应中间体,化学反应产物的产率取决于地磁场相对于反应物分子的方向[6]。
乍一看,这种理论几乎难以置信:在生理条件下,地球磁场(30-65 μT)与单个分子的相互作用比其热能k B T小(xiǎo)一百万倍以上,很难对热化学反应动力学或热力学产生影响。然而,不妨设想这样一个场景:有一个沉重的石块,苍蝇是否能够通过撞击来使其倾倒?答案显然是否定的。但是,如果(guǒ)先(xiān)给(gěi)予(yǔ)一(yī)定(dìng)的(de)能(néng)量(liàng),使(shǐ)石(shí)头(tóu)仅(jǐn)靠(kào)一(yī)条(tiáo)棱(léng)保持平衡。此时即使苍蝇撞击传递的能量很小,也足以导致块落到右侧而不是左侧。
图四 自由基对机制的类比模型[7]
在化学反应热力学系统中,平衡态均处于能量谷(gǔ),不(bù)同(tóng)之(zhī)处(chù)在(zài)于(yú)前(qián)者(zhě)是(shì)稳(wěn)定(dìng)平(píng)衡(héng),位(wèi)于(yú)能(néng)量(liàng)最(zuì)低(dī)的(de)谷(gǔ);后(hòu)者(zhě)则(zé)属(shǔ)于(yú)不(bù)稳(wěn)定(dìng)平(píng)衡(héng),能(néng)量(liàng)谷(gǔ)的(de)相(xiāng)对(duì)值(zhí)高(gāo)于(yú)反(fǎn)应(yīng)物(wù)和(hé)产(chǎn)物(wù),微(wēi)小(xiǎo)的(de)扰(rǎo)动(dòng)即(jí)可(kě)使(shǐ)系(xì)统(tǒng)偏(piān)离(lí)本(běn)来(lái)状(zhuàng)态(tài),跌(diē)至(zhì)能(néng)量(liàng)更(gèng)低(dī)的(de)另(lìng)一(yī)个(gè)谷(gǔ)。自(zì)由(yóu)基(jī)对(duì)机(jī)制就(jiù)是(shì)利(lì)用(yòng)非(fēi)平(píng)衡(héng)态(tài)系(xì)统(tǒng)的(de)不(bù)稳(wěn)定(dìng)性(xìng)发(fā)生(shēng)的(de)反(fǎn)应(yīng):非(fēi)平(píng)衡(héng)态(tài)是(shì)自(zì)由(yóu)基(jī)对(duì),达(dá)到(dào)该(gāi)状(zhuàng)态(tài)所(suǒ)需(xū)的(de)能(néng)量(liàng)来(lái)自(zì)光(guāng)子(zi)。
那么,反应的最佳载体是什么呢?常规的哺乳动物光感受器视蛋白将光能转化为一种视觉信号转导分子(视黄醛)的构象变化,不涉及自由基对反应;然而对鸟类视觉系统的研究表明,候鸟的眼睛中还存在隐花色素,且其种类多达四种[8],具有发生自由(yóu)基(jī)对(duì)反(fǎn)应(yīng)的(de)潜(qián)力(lì)。
接(jiē)下(xià)来(lái),我(wǒ)们(men)就(jiù)简(jiǎn)要(yào)介(jiè)绍(shào)一(yī)下(xià)自(zì)由(yóu)基(jī)对(duì)反(fǎn)应(yīng)机(jī)理(lǐ)及(jí)其(qí)实(shí)现(xiàn)磁(cí)场(chǎng)感(gǎn)知(zhī)的(de)机(jī)制(zhì),并探讨隐花色素是如何介导自由基反应发生的。
一、自由基对反应机理
我们知道物质由原子组成,原子又由原子核围绕电子形成。稳定状态下,电子总是以两两成对的形式存在,但化学反应中有可能产生包含未成对电子的物质,也就是自由基。一个自由基对由两个自由基组成,这两个自由基通常是通过单分子均裂或者双分子间电子转移产生的。例如甲烷(CH4)总共有10个电子,其中 6 个由碳贡献,4个来自氢。两个内层电子围绕着碳原子核,八个外层电子参与形成碳氢键,每个键两个电子。如果其中一个键均裂,得到两个电中性基团:甲基自由基 CH3 • 和氢原子H•。
由于电子具有自旋角动量的特性,类似一个绕其轴旋转的小球形物体,因此当电子带电旋转,会产生一个磁场,就像线圈中的电流,因此电子可视作一个微观磁铁,具有一个固定磁矩。CH3 • 有 9 个电子,其中 8 个成对存在,因此它们的磁矩正好抵消。因此,自由基的磁性源于未成对电子。
电子自旋角动量的特殊之处在于其具有量子特性,大小由自旋量子数 S 决定,单位为±1/2。量子力学规定,一个自由基对如CH3 • 和H• 的自旋量子数只能为 0 或 1,可以理解为两个不成对电子的自旋可以平行(↑↑, S=1)或反平行(↑↓,S=0)。由于理论认为自旋量子数为S的物质有2S+1种态,故S = 1为三重态,S=0为单重态;因此,自(zì)由(yóu)基(jī)对(duì)的(de)两(liǎng)种(zhǒng)形(xíng)式(shì)称(chēng)为(wèi)三(sān)元(yuán)组(zǔ)(S=1) 和(hé)单元组(S=0)。
自由基对反应中,电子必须自旋守恒。由于自由基反应的产物为单元组的分子,因此最直接的中间体自由基必须也为单元组,而三元组自由基则由单元组自由基转化而来。
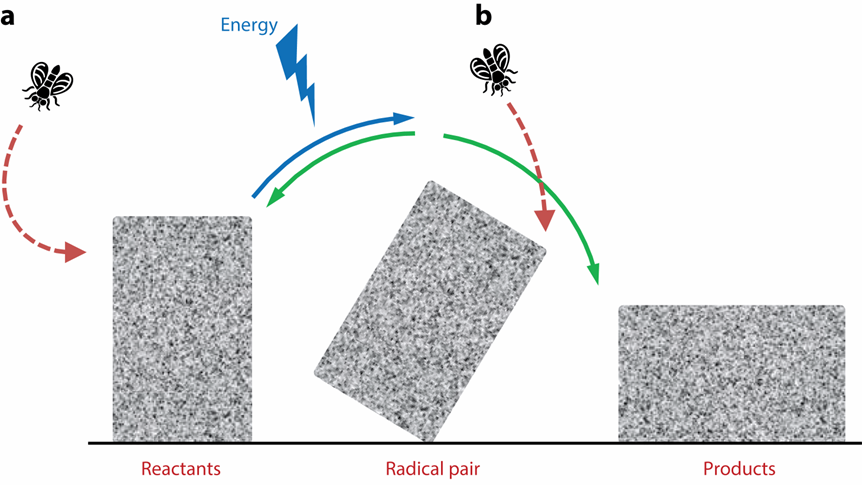
此外,自由基对还具有两个(gè)显(xiǎn)著(zhe)的(de)特(tè)征(zhēng):其(qí)一(yī),存(cún)在(zài)广(guǎng)泛(fàn)的(de)超(chāo)精(jīng)细(xì)相(xiāng)互(hù)作(zuò)用(yòng),可(kě)以(yǐ)简(jiǎn)单(dān)理(lǐ)解(jiě)为(wèi)自(zì)由(yóu)基(jī)中(zhōng)离(lí)域的(de)不(bù)成(chéng)对(duì)电(diàn)子(zi)同(tóng)时(shí)与(yǔ)多(duō)个(gè)原(yuán)子(zi)核(hé)相(xiāng)互作用,且这种作用是不对(duì)称(chēng)的(de)(随角度变化),构成了磁场方向感知的基础;其二,由于量子力学原因,单重态和三重态之间存在高频相(xiāng)互(hù)转(zhuǎn)化(huà),也(yě)即(jí)单(dān)重(zhòng)态(tài)分(fēn)子(zi)的(de)占(zhàn)比(bǐ)处(chù)于(yú)高(gāo)频(pín)振(zhèn)荡中(图五a)。这一震荡是由内部原子核自旋引起的,也因此,这一震荡很容易受到外部磁场波形的调制:不同频率的波叠加,形成低频包络波形和高频振荡波形。而由于超精细相互作用的不对称性,不同方向的磁场会产生不同的叠(dié)加(jiā)波(bō)形(xíng)(图(tú)五(wǔ)b、c)。
图(tú)五(wǔ) 单(dān)重(zhòng)态(tài)自(zì)由(yóu)基(jī)分(fēn)数(shù)随(suí)时(shí)间(jiān)变(biàn)化(huà)情(qíng)况(kuàng)[7]
基(jī)于(yú)上(shàng)述(shù)的(de)自(zì)由(yóu)基(jī)对(duì)反(fǎn)应(yīng)特(tè)征(zhēng),一(yī)个(gè)精(jīng)致(zhì)的(de)模(mó)型(xíng)应(yīng)运(yùn)而(ér)生(shēng)(图(tú)六(liù)a):光(guāng)激(jī)发(fā)下(xià),一(yī)个(gè)单(dān)重(zhòng)态(tài)自(zì)由(yóu)基(jī)对(duì)瞬(shùn)时(shí)形(xíng)成(chéng),成(chéng)为(wèi)两(liǎng)个(gè)相(xiāng)互(hù)竞(jìng)争(zhēng)的(de)反(fǎn)应(yīng)的(de)原(yuán)料(liào):其(qí)可(kě)以(yǐ)直(zhí)接(jiē)或(huò)间(jiān)接(jiē)(转(zhuǎn)化(huà)为(wèi)三(sān)重(zhòng)态(tài))生(shēng)成(chéng)产(chǎn)物(wù),也(yě)可(kě)以(yǐ)通(tōng)过(guò)逆(nì)反(fǎn)应(yīng)回(huí)到反应物。
如果与外场的相互作用增加了平均三重态分数(单重态分数相应减少),那么更少的自由基对将恢复回 AB,更多的自由基对生成 C 做出贡献,反之亦然。因此C 的最终产率取决于外部磁场的存在和方向。这就是磁场效应的起源。尽管自由基对振荡对于磁场效应的存在至关重要,但一旦所有自由基对都消失,C 的最终产率将为鸟类提供有关磁场方向的信息。
这个模型可以做如下类比(图六b):两个标有 S 和 T 的浴缸由一根带阀门的管子连接,其中的水量分别对应于单重态和三重态自由基对的分数。T 在底部有一个出口,该出口排空到标记为 C 的桶中,而 S 有两个出口,其中一个通向 C,另一个通向AB。初始情况, S 装满了水,T、AB、C 为空,瓣膜部分打开。水从 S 流到 T,同时落入桶中,直到浴缸排空。最终进入两个桶中的水量(AB 和(hé) C 的最终产量)取决于管子的直径(速率常数k S 和 k C 的值)以及将阀门转动的程度(外加磁场对转化(huà)情(qíng)况(kuàng)的(de)调(diào)制(zhì))。如(rú)果(guǒ)阀(fá)门(mén)完(wán)全打(dǎ)开(kāi),使(shǐ)水(shuǐ)从(cóng) S 快(kuài)速(sù)流(liú)向(xiàng) T,则(zé)有(yǒu)两(liǎng)条(tiáo)有(yǒu)效(xiào)的(de)路径到(dào)达(dá) C;如(rú)果(guǒ)阀(fá)门(mén)部(bù)分(fēn)关闭(bì),则(zé)到(dào)达(dá) T 的(de)水(shuǐ)就(jiù)会(huì)减(jiǎn)少(shǎo),到(dào)达(dá) C 的(de)唯(wéi)一有效路线是直接从 S 到达,最后C 中的水比阀门完全打开时少。
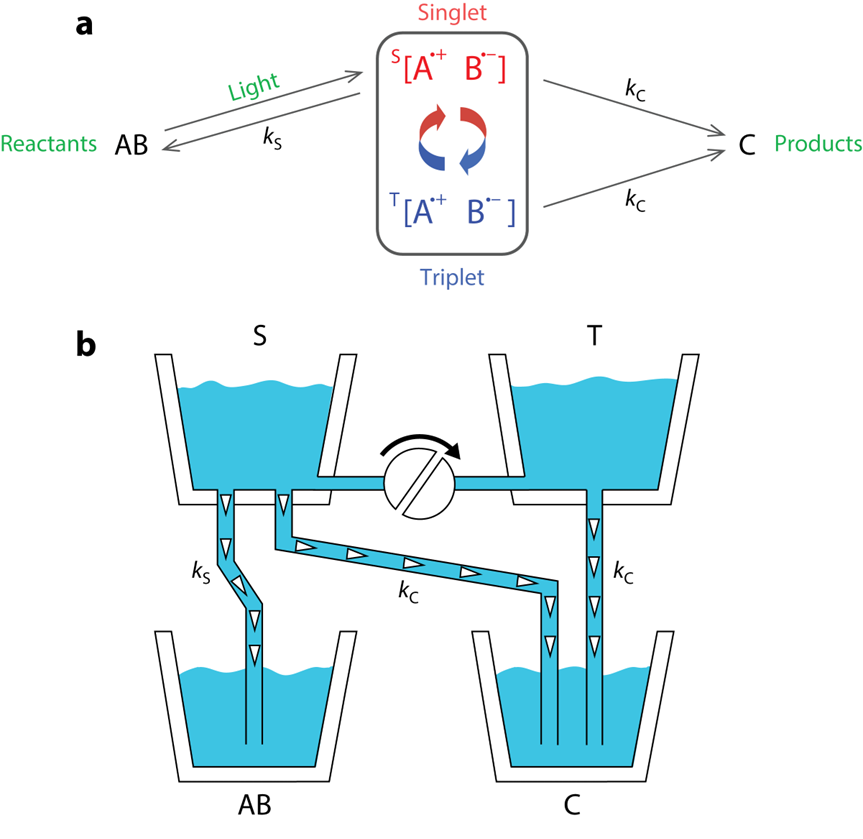
图六 自由基对反应机理示意图[7]
二、隐花(huā)色(sè)素(sù)参(cān)与(yǔ)磁(cí)场(chǎng)方(fāng)向感知
隐花色素,顾名思义,是最早在植物(wù)中(zhōng)发(fā)现的一种感光色素,参与植物生长发育的光依赖性调节。其有一个结合域非共价结合FAD(氧化还原活性黄素腺嘌呤二核苷酸辅因子,其还原态FADH参与呼吸作用电子传递链的重要一环,其氧化态FAD则可以吸收蓝光)。FAD 的光还原是通过色氨酸(Trp,W) 残基三联体的三个连续电子转移(yí)介(jiè)导的,生成黄酮半醌自由基 FAD−•和TrpH+•(图七a)。
进一步的生化实验证实了隐花色素参与自由基对反应机制,自由基对中间体命名为RP1(radical pair1),而产物C对应次级自由基,命名为RP2(图七b)。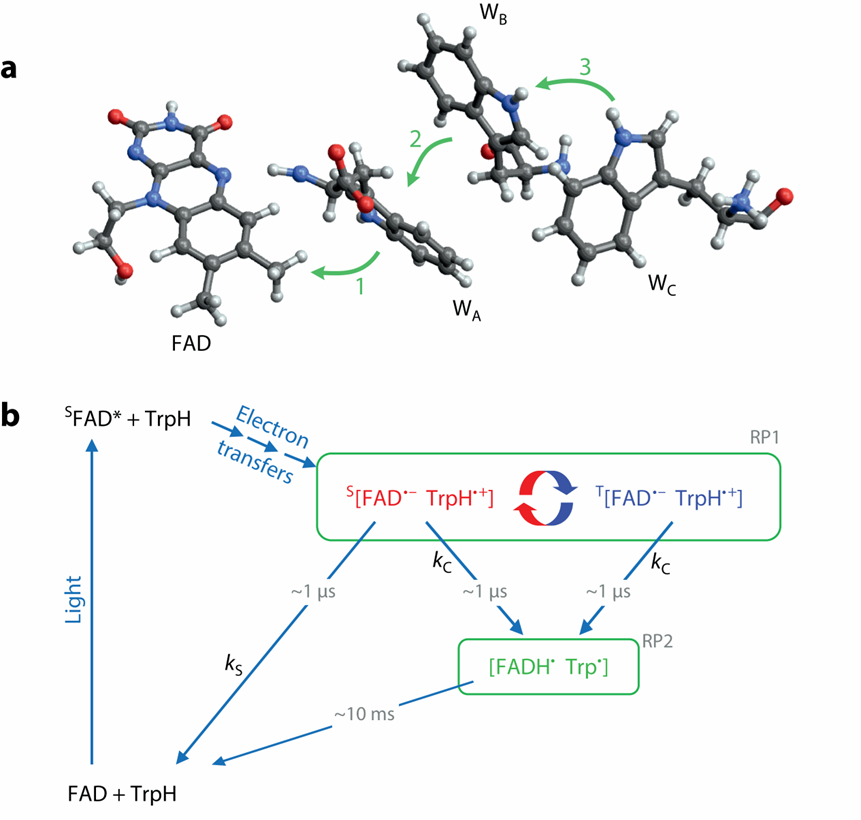
图(tú)七(qī) 隐(yǐn)花(huā)色(sè)素(sù)参(cān)与(yǔ)自(zì)由(yóu)基(jī)对(duì)反(fǎn)应(yīng)的(de)化(huà)学(xué)机(jī)理(lǐ)[7]
至(zhì)此(cǐ),化(huà)学(xué)家(jiā)证(zhèng)明(míng)了(le)自(zì)由(yóu)基(jī)对(duì)反(fǎn)应(yīng)介(jiè)导(dǎo)磁(cí)场(chǎng)方(fāng)向(xiàng)感(gǎn)应(yīng)的(de)可(kě)能(néng)性(xìng),并(bìng)证(zhèng)明隐(yǐn)花(huā)色(sè)素(sù)是(shì)一(yī)种(zhǒng)很(hěn)有(yǒu)希(xī)望(wàng)的(de)候(hou)选(xuǎn)分(fēn)子(zi)。然(rán)而(ér)事(shì)实(shí)是(shì)否(fǒu)如(rú)此(cǐ)?这(zhè)个(gè)问(wèn)题(tí)需(xū)要(yào)由(yóu)生(shēng)物(wù)学(xué)家(jiā)回(huí)答(dá)。
有(yǒu)研(yán)究(jiū)表(biǎo)明(míng),在(zài)夜(yè)间(jiān)活(huó)跃(yuè)的(de)候(hou)鸟(niǎo)中(zhōng),磁(cí)取(qǔ)向(xiàng)行(xíng)为(wèi)期(qī)间(jiān)的(de)神(shén)经(jīng)活(huó)动(dòng)与(yǔ)视(shì)网(wǎng)膜(mó)神(shén)经(jīng)节(jié)细(xì)胞(bāo)中(zhōng)的(de)隐(yǐn)花(huā)色(sè)素(sù)表(biǎo)达(dá)共(gòng)定(dìng)位(wèi),而(ér)在(zài)非(fēi)候(hou)鸟(niǎo)或(huò)白(bái)天(tiān)没(méi)有(yǒu)这(zhè)种(zhǒng)共(gòng)定(dìng)位(wèi)[9]。遗(yí)憾(hàn)的(de)是(shì),目(mù)前(qián)尚(shàng)缺(quē)乏(fá)更(gèng)为(wèi)直(zhí)接(jiē)的(de)证(zhèng)据(jù),只(zhǐ)能(néng)证(zhèng)明(míng)隐(yǐn)花(huā)色(sè)素(sù)参(cān)与(yǔ)磁(cí)场(chǎng)感(gǎn)知(zhī),但(dàn)不(bù)能(néng)排(pái)除(chú)它(tā)在(zài)磁(cí)场(chǎng)感(gǎn)知(zhī)的(de)上(shàng)游(yóu)或(huò)下(xià)游(yóu)起(qǐ)间(jiān)接(jiē)作(zuò)用(yòng)的(de)可(kě)能(néng)性(xìng)。此(cǐ)外(wài),行(xíng)为(wèi)学(xué)实(shí)验(yàn)发(fā)现(xiàn)磁(cí)感(gǎn)受(shòu)的(de)机(jī)制(zhì)只(zhǐ)存(cún)在(zài)于(yú)右(yòu)眼(yǎn):被(bèi)测(cè)试(shì)的(de)迁(qiān)徙(xǐ)鸟(niǎo)类(lèi)被(bèi)遮(zhē)住(zhù)左(zuǒ)眼(yǎn)时(shí),定(dìng)向(xiàng)能(néng)力(lì)并(bìng)未(wèi)出(chū)现(xiàn)显(xiǎn)著(zhe)下(xià)降(jiàng),而(ér)被(bèi)遮(zhē)住(zhù)右(yòu)眼(yǎn)时(shí)未(wèi)能(néng)表(biǎo)现(xiàn)出(chū)定(dìng)向(xiàng)行(xíng)为(wèi)(图(tú)十(shí)二(èr))[10]。未来需要更多操纵性的实验建立起从分子、细胞到环路和行为的系统研究。
看似微不足道的磁场能量,通过感光色素的自由基反应巧妙调控了物质的产率,以千分之一秒的精度在鸟类的视网膜上书写精确而具体的磁场信息——行文至此,不知你是否和我一样,沉思并且赞叹,感慨自然选择的神奇和时间的强大。
仰望天穹,碧蓝的寰宇衬托出飞鸟纤细的身姿,它们眼中是否有一条条磁力线绵延而过?或许化学家和生物学家永远不能回答这个问题;但至少在某一点上,我们和千年前的古人达成了共识——远航的终点,是家的方向。
参考文献:
[1]Mouritsen, Henrik. “Long-distance navigation and magnetoreception in migratory animals.” Nature vol. 558,7708 (2018): 50-59. doi:10.1038/s41586-018-0176-1
[2]Johnsen, Sönke, and Kenneth J Lohmann. “The physics and neurobiology of magnetoreception.” Nature reviews. Neuroscience vol. 6,9 (2005): 703-12. doi:10.1038/nrn1745
[3]von Middendorff A. Die Isepiptesen Rußlands. Mém Acad Sc. S Petersbourg VI Ser Tome. 1859;8:1–143.
[4]Wiltschko, W. (1968), Über den Einfluß statischer Magnetfelder auf die Zugorientierung der Rotkehlchen (Erithacus rubecula). Zeitschrift für Tierpsychologie, 25: 537-558. https://doi.org/10.1111/j.1439-0310.1968.tb00028.x
[5]Wiltschko, W., Wiltschko, R. Disorientation of inexperienced young pigeons after transportation in total darkness. Nature 291, 433–434 (1981). https://doi.org/10.1038/291433a0
[6]Schulten, K., Swenberg, C. & Weller, A. (1978). A Biomagnetic Sensory Mechanism Based on Magnetic Field Modulated Coherent Electron Spin Motion. Zeitschrift für Physikalische Chemie, 111(1), 1-5. https://doi.org/10.1524/zpch.1978.111.1.001
[7]Hore, P J, and Henrik Mouritsen. “The Radical-Pair Mechanism of Magnetoreception.” Annual review of biophysics vol. 45 (2016): 299-344. doi:10.1146/annurev-biophys-032116-094545
[8]Nießner C, Gross JC, Denzau S, Peichl L, Fleissner G. 149. et al. 2016. Seasonally changing cryptochrome 1b expression in the retinal ganglion cells of a migrating passerine bird. PLOS ONE 11:e0150377
[9]H. Mouritsen, U. Janssen-Bienhold, M. Liedvogel, G. Feenders, J. Stalleicken, P. Dirks, R. Weiler, Cryptochromes and neuronal-activity markers colocalize in the retina of migratory birds during magnetic orientation, Proc. Natl. Acad. Sci. U.S.A.101 (39) 14294-14299,https://doi.org/10.1073/pnas.0405968101 (2004).
[10]Wiltschko, Wolfgang et al. “Lateralization of magnetic compass orientation in a migratory bird.” Nature vol. 419,6906 (2002): 467-70. doi:10.1038/nature00958